|
Vstup pro předplatitele: |
VERONICA - ČASOPIS PRO OCHRANU PŘÍRODY A KRAJINY




Poslední čísla
 | |
|  |
 |  |

Obr. 2 - Kořen s gelovou vrstvou mucilagu, tedy jakéhosi lepivého slizu (za kořenovou čepičkou), proniká mezi půdní agregáty a je kolonizován podpůrnými mikroorganismy. Mucilag je vylučován rostlinou pro ochranu citlivých, čerstvě vznikajících pokožkových buněk na povrchu tloustnoucí části kořene. Z vrcholové části kořene jsou ale také v podobě nízkomolekulárních sloučenin uvolňovány informace o pronikání kořene konkrétní rostliny, o konci „hladového období“ v půdě. Mucilag představuje totiž mimo jiné stimulační organické hnojení podporující aktivitu podpůrných půdních mikroorganismů, také ale bariéru proti infekci patogenními mikroorganismy.

Obr. 3 - Výměna informací jednak mezi bakteriemi navzájem a také mezi bakteriemi a rostlinou je velmi složitá. Půdní bakterie mají nejreaktivnější části svých buněk, povrchové biomembrány, vystaveny účinkům nepřeberného množství půdních látek. Aby byly úspěšné, musí reagovat okamžitě nebo vůbec. Proto neustále sledují okolí a prostřednictvím vnitrodruhových (světle zabarvených) a mezidruhových (tmavě zabarvených) signálních látek vylučovaných bílkovinnými kanálky a přijímaných příslušnými receptory koordinují své chování. Vzory chování, zda budou dále využívat individuální, úsporné strategie, anebo přejdou na spontánní kolektivní strategie (pevné šipky), nastávají při zjištění změny početnosti buněk. K regulaci chování bakterií dochází také na základě příjmu napodobenin signálních látek, které jsou vylučovány rostlinou (světlá barva signálních látek vylučovaných kořenem vpravo nahoře).

Obr. 4 - Stimulace kolonizujících mikroorganismů kořenovými výměšky (exsudáty) vede automaticky ke zvýšeným nárokům na výživu rychle se množících mikrobních společenstev. Zásoby dostupných živin v okolním půdním roztoku jsou velmi rychle vyčerpány zabudováním do buněk mikroorganismů. Zvyšuje se druhová pestrost, biodiverzita mikroorganismů. Mikroorganismy dlouho nestrádají. Signální kaskády mikroorganismů spouští vylučování vhodných mimobuněčných – extracelulárních enzymů (označené hvězdičkami [ECE]), které jsou schopny uvolnit nedostatkové živiny z alternativních zdrojů, z půdních agregátů. Živiny se zde nacházejí v tmelech nebo povlacích na minerálních částicích půdy. Jsou navázané ve formě částečně rozložených látek, které zůstaly zachovány z předcházejících vegetačních sezon po interakcích mezi rostlinami a půdními organismy a půdou. Před pokračujícím mikrobiálním rozkladem byly dosud chráněny různými mechanismy. V podmínkách nadbytku energetických zdrojů a nedostatku živin se ale situace mění, jsou vytvořeny příznivé podmínky pro další rozklad, pro uvolnění živin v redukovaných nebo oxidovaných minerálních formách (Nred, Nox, Pox, Sox), které jsou horlivě přijímány (kontrolovány) mikroorganismy. Nároky rostlin na živiny nejsou v tomto stadiu ještě vyslyšeny. Kořeny „ovládají“ rhizosféru dodáváním uhlíkatých látek bohatých na energii, mikrobní společenstva „ovládají“ rhizosféru biochemicky produkcí klíčových enzymů pro zpřístupnění živin.
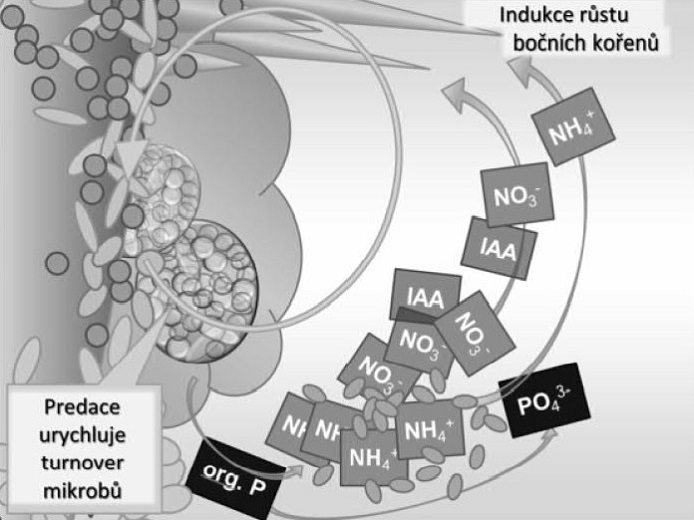
Obr. 5 - Velké množství mikrobiální biomasy láká půdní predátory, v našem případě půdního prvoka, pro kterého jsou buňky bakterií svým prvkovým a látkovým složením velmi blízké a snadno stravitelné. V mikrobní biomase ale konzumenti nedostávají dostatečné množství energeticky bohatých látek, např. sacharidů, což vyvažují zvýšením množství přijímané mikrobiální biomasy. Nadbytečně přijímané živiny, např. dusík a fosfor, vylučují predátoři v exkrementech. Konzumace bakterií paradoxně obnovuje následnou vysokou aktivitu přežívajících mikroorganismů. Není-li amonný dusík odebírán rostlinou či mikroorganismy, je dále transformován nitrifikačními bakteriemi na dusík nitrátový, který se v půdě snadno pohybuje. Malý koloběh dusíku se uzavírá. Za „uhlíkaté investice“ do půdy získává rostlina odpovídající množství cenných živin včetně biologicky aktivních stimulačních látek, rostlinných hormonů (IAA). A ani půda nestrádá, má pro své „děti“ více energie uložené do chemických vazeb, než měla na začátku.
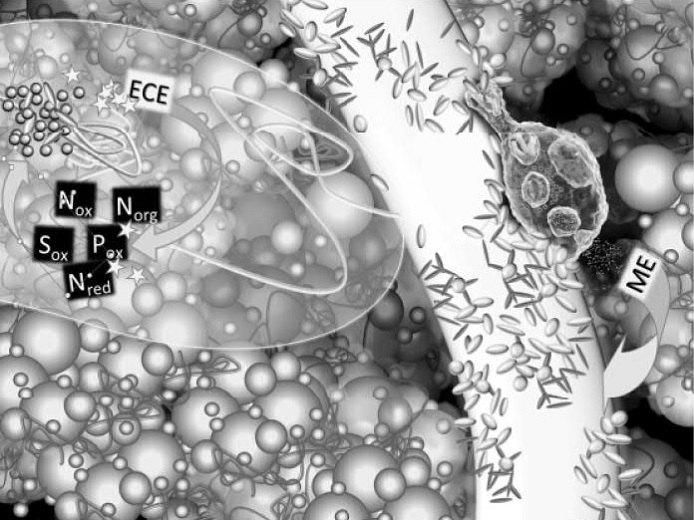
Obr. 6 - Výrazné limitování živinami v okolí kořenů ve srovnání s okolní půdou vede k intenzivnímu soutěžení o živiny mezi kořeny a mikroorganismy. Navzdory této soutěži jsou mikroorganismy a kořeny na sobě závislé a vyvinuly různé formy symbióz. Jednou z nich je infikování rostlinného kořene spolupracující mykorhizní houbou, jejíž houbová vlákna umožňují transport živin ze vzdálenějších míst v půdě (část půdy ovlivněná mykorhizní houbou – na obrázku je vyznačena nalevo od kořene ohraničenou průhlednou zónou). Další výhodou je schopnost houbových vláken pronikat i do menších pórů, do jakých by nepronikly ani ty nejjemnější kořeny. Rostlinám je umožněno přenést přes houbová vlákna vlastní investice atraktivních uhlíkatých látek do podstatně většího objemu půdy. V těchto místech potom dochází ke stejným reakcím mezi kořenem, mikroorganismy a půdou, jaké již známe z nejbližšího okolí kořene (viz obr. 4). Signální kaskády mikroorganismů a mykorhizních hub spouští vylučování extracelulárních enzymů (označené hvězdičkami [ECE]), které jsou schopny uvolnit nedostatkové živiny ze vzdálenějších půdních agregátů. Výsledkem je uvolnění živin v minerálních formách (Nred, Nox, Pox, Sox), které jsou horlivě přijímány houbami a mikroorganismy. Mykorhizní symbióza umožňuje pokrýt až 90 % rostlinných nároků na fosfor a 80 % nároků na dusík. Kromě toho získává rostlina soužitím s houbou také výhodu indukované ochrany před určitými skupinami houbových patogenů. Explozivní nárůst počtu mikroorganismů ale netrvá dlouho, pouze v měřítku hodin až dnů. Rychle narůstající mikrobiální biomasa láká konzumenty mikrobů, zástupce půdní fauny, predátory, zejména z řad půdních prvoků a hlístic, pro které jsou mikrobní buňky s obdobným látkovým složením velmi atraktivním zdrojem výživy. Modelový prvok je znázorněn vpravo nahoře na povrchu kořene, zřetelně je vidět příjem potravy buněčnými ústy, dále bakterie v potravních vakuolách a vylučování buněčnou řití. Vylučované látky jsou bohaté na jednoduché sloučeniny životodárných prvků, dusíku, fosforu, síry apod. Koloběh se uzavírá, takto uvolněné živiny [ME] již rostlinný kořen přijímá.
Doporučujeme ke čtení
Zpracovávám rád každé zadání, protože nejsou nezajímavá témata…
Pavel Dvorský, č. 1/2025, str. 23, pro předplatiteleKšefty nám bezostyšně ničí přírodu a krajinu
Hynek Skořepa, č. 1/2025, str. 32Jurkovičovy válečné hřbitovy
Václav Štěpánek, č. 1/2025, str. 7, pro předplatitelePtačí lokalita Ústřední hřbitov v Brně
Karel Hudec († 10. 11. 2017), č. 1/2025, str. 14, pro předplatiteleTřikrát pár slov k Tragikomedii ochrany přírody Tomáše Grima
č. 1/2025, str. 36Miesta stretnutí
Denisa Halajová, č. 1/2025, str. 16, pro předplatiteleStaré cintoríny ako súčasť historických štruktúr krajiny
Mikuláš Huba, č. 1/2025, str. 2, pro předplatiteleTragikomedie ochrany přírody
Tomáš Grim, č. 4/2024, s. 2-5Třikrát pár slov k Tragikomedii ochrany přírody Tomáše Grima
č. 1/2025, str. 36Dostat pořádný kartáč není na škodu
Pavel Pechoušek, Vilém Jurek, č. 4/2024, s. 10-12Reportáž opožděná o třicet pět let
Václav Štěpánek, č. 4/2024, s. 34-37Odkaz Josefa Vavrouška je dnes dvojnásobne aktuálny
Mikuláš Huba, č. 3/2024, s. 28-31Nová chráněná území v Brně
Vilém Jurek, č. 2/2024, s. 32-33



Interakce rostlin, mikroorganismů a půdy
Jaroslav Záhora, č. 1/2018, s. 6-9
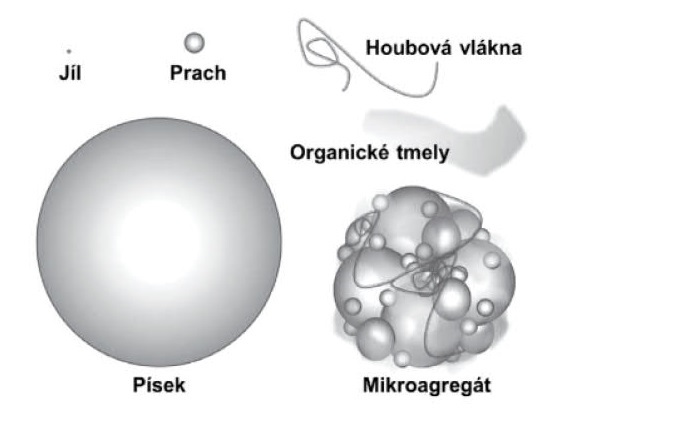
Obr. 1 - Menší půdní částice půdotvorného materiálu, pojmenované podle velikosti jíl, prach a písek, se shlukují do různě velkých hrudek, do půdních agregátů. Jsou spojené organickými či organominerálními tmely. Podle velikosti je lze rozdělit na mikroagregáty (menší než 250 μm) a makroagregáty (větší než 250 μm). Schopnost půdy shlukovat se určuje půdní strukturu. Relativní zastoupení jílu, prachu, písku a skeletu určuje texturu půdy, kterou charakterizujeme půdní druhy; půdy jílovité, hlinité, písčité, příp. kamenité (více než 20 % skeletu). Pro zemědělství jsou nejvhodnější půdy hlinité (středně těžké), ve kterých převažují částice o velikosti 0,01–0,1 mm a které jsou dobře propustné pro vodu i vzduch.
Půda je jedinečná hmota zajišťující podmínky pro život na Zemi. Pouhé čtyři složky, pevná, kapalná, plynná a živá, vytvářejí nejsložitější oživený systém na planetě. Klíčová je přitom složka živá nacházející se v dalších dvou složkách půdy, v organické a minerální. Vše je výsledkem dlouhodobého vývoje. Z ekosystémového hlediska zajišťuje půda nejen produkci potravin, krmiv a vlákniny, ale prostřednictvím ukládání uhlíkatých látek také regulaci klimatu. Půda je určující složkou ekosystému. Zásadním způsobem ovlivňuje koloběhy vody a biogenních prvků. Dochází v ní k přirozenému čištění prosakujících vod a ke snižování obsahu cizorodých látek. Smutné je, že současné tempo ztrát a zhoršování kvality půd je rychlejší, než je doba nutná na regeneraci půdy nebo na tvorbu půdy nové. Odhaduje se, že 1 cm půdy vzniká průměrně za dobu 200 let. To vše umocňuje význam půdy jako konečného zdroje.
Půdní agregáty, tedy základní půdní strukturní jednotky uspořádání pevných částic, jsou navazujícím uspořádáním původních minerálních částic, které je obohaceno o organominerální pojiva (oxidy, humusotvorné látky, mikrobiální biofilmy, vlákna půdních hub, kořeny rostlin s jejich výměšky apod.). Půdní agregáty ovlivňují velikost a množství pórů, které jsou vyplněné vzduchem nebo vodou. Shlukování půdních částic do mikroagregátů a jejich spojování do větších agregačních celků zvětšuje infiltrační, tedy vsakovací, a retenční neboli zadržovací schopnost půdy. Zvětšuje množství srážkové vody, kterou je půda schopna přijmout a zadržet. Uvnitř půdních agregátů je navíc velmi dobře „hlídáno“ to nejcennější půdní bohatství; nejmenší půdní organismy, organické látky a živiny. Čím větší půdní agregáty totiž půda má, tím víc je přímý kontakt mezi kořenem a půdou omezen a rostliny jsou ve větší míře „odkázány“ na spolupráci s mikroorganismy. Na získání odpovídajícího množství živin, na „vnitropůdní obchodování“, musí rostliny vynaložit více uhlíkatých investic, více uhlohydrátů bohatých na energii, a to i za cenu dílčího omezení nadzemní produkce. Tím je zajišťována blahodárná regenerace organické hmoty stejně tak jako infiltrační a retenční schopnost půdy.
Povrch rostlinného kořene je možná nejzajímavějším místem kontaktů mezi živou a neživou přírodou. Je místem, kde se rozhoduje o tom, jak bujná a druhově bohatá bude vegetace, jak velká část látek získaných při fotosyntéze bude rozdělena mezi nadzemní a podzemní biomasu rostlin, jak budou přítomné organismy vzájemně závislé, dokonce i o tom, jak snadno bude ekosystém otevřený nepůvodním druhům. V blízkosti kořene je možno najít pokornou obhajobu organického přístupu k zemědělství.
Tím, kdo rozpoznal význam zvýšených aktivit půdních organismů spojených s přítomností kořenového systému a kdo již v roce 1904 navrhl pro oblast vlivu rostlinných kořenů v půdě označení „rhizosféra“, byl Lorenz Hiltner, ředitel Bavorského zemědělsko-botanického ústavu v Mnichově. Jeho výzkumy z oblasti půdní bakteriologie a rhizosféry natrvalo ovlivnily chápání vzájemného ovlivňování rostlin a půdních organismů. Přesto vědci dlouho přehlíželi biologickou podstatu výměny látek a energií na rozhraní mezi půdou a rostlinou. Úloze kořenů při získávání živin a vody byla zpočátku věnována jen malá pozornost. Převládajícím názorem byla minerální výživa rostlin. Jsou dokonce státy, kde takový stav trvá dodnes.
Po více než sto letech jsou nejnovějšími vědeckými metodami potvrzovány Hiltnerovy závěry o významných změnách v rhizosféře, tedy o změnách, které si rostliny samotné řídí uvolňováním uhlíkatých sloučenin. Nabídka pestré škály atraktivních organických látek v okolí kořínků označovaná jako rhizodepozice, nepříliš přesně výměšky kořenů, představuje souhrnně 20-50 % (výjimečně až 80 %) z primární produkce rostlin. Intenzita biochemických dějů v rhizosféře vyvolaná těmito kořenovými depozicemi dosahuje o několik řádů vyšších hodnot než v okolní půdě. Rhizosféra může obsahovat od 106 až do 109 bakterií, 104 prvoků, od 101 až po 102 hlístic a od 105 po 106 hub na gram půdy.
V nerhizosférní půdě, v půdě neovlivněné přítomností kořenů, mikroorganismy strádají a přežívají s minimálními aktivitami, přičemž tak nedobrovolně chrání cennou neúplně rozloženou část půdní organické hmoty. Zjevné „strádání“ půdních mikroorganismů bylo empiricky prokázáno již v sedmdesátých letech 20. století na základě rozporu mezi krátkou generační dobou mikroorganismů (doba nutná pro zdvojnásobení počtu mikroorganismů v populaci) naměřenou v optimálních podmínkách laboratoří (cca 20 hod.) a dobou jejich životnosti v půdě (1-1,5 roku). Vlastní „strádání“, související s efektivním využíváním organických látek a se zachováním přirozené půdní úrodnosti, není vyvoláno pouze nedostatkem klíčových biogenních prvků, ale souhrou mezi více činiteli. Podstatný vliv mají fyzikální, chemické a biologické charakteristiky půdy, konkurenční vazby mezi půdními mikroorganismy, složitosti potravních sítí a vazeb, koncentrace signálních látek apod. V rhizosférní půdě se z kořene uvolňují bohaté a rozmanité zdroje uhlíkatých látek a energie, čímž je stimulována bouřlivá aktivita půdních mikroorganismů, z níž mají prospěch půdní bezobratlí i rostlina.
„Paradox Šípkové Růženky“
Životodárný, „resuscitační“ účinek kořenových výměšků na bouřlivé aktivity mikroorganismů a následně půdních bezobratlých lze přirovnat k polibku prince z pohádky o Šípkové Růžence. V roce 1995 využívá prof. Lavelle popisovaného příměru označeného jako „paradox Šípkové Růženky“ („Sleeping Beauty paradox“) pro popis přechodu mikroorganismů ze stadia kolektivního klidového období se snížením metabolismu do stadia extrémních aktivit.
Bouřlivé aktivity organismů na rozhraní rostlinného kořene a půdy jsou vyvolány jejich snahou získat základní prvky pro syntézu vlastních buněk ve výjimečném prostředí nadbytku uhlíkatých látek a energie. Všechny živé organismy vyžadují totiž pro udržení vlastní životaschopnosti, pro růst a pro reprodukci téměř stejné živiny. Tento jednotící zájem je základem pro soutěžení mezi rostlinami a mikroorganismy v půdách s konečnou nabídkou živin. Zde nevystačíme s obecným konstatováním, že rostlina ovládá půdní prostředí uhlíkově a energeticky a mikroorganismy biochemicky, i když není na škodu si to připomenout.
Nepřetržitý odběr živin rostlinami vede ke vzniku silně vyčerpaných zón kolem kořenů. V této zóně, cca 1-3 mm od povrchu kořene, se koncentrace živin, které se doplňují pomalu z okolní půdy, jako je např. fosfor, může snížit cca 5-10krát ve srovnání s okolním půdním prostředím. Na druhé straně, mobilní formy živin v půdním roztoku, např. NO3-, kolem kořenů vyčerpané zóny nevytvářejí.
Konzumace mikroorganismů na povrchu kořene je přitom pro výživu rostlin životně důležitá. Žírem mikroorganismů predátoři paradoxně obnovují jejich aktivitu. Znovu se zrychlují látkové výměny mezi rostlinou, půdou a mikrobiálními společenstvy. Je tu ale ještě jeden háček. V mikrobní biomase nemají konzumenti dostatek energie, tedy dostatečné množství energeticky bohatých látek, např. sacharidů. „Dietetickou chybu“ nedostatku energie v jinak atraktivní potravě tvořené buňkami mikrobů vyvažují predátoři zvýšením množství přijímané biomasy. V takovém okamžiku se ale stává přijímaný mikrobní dusík, fosfor a síra pro predátory nadbytečnou přítěží, kterou vylučují v exkrementech. A právě na tuto chvíli, na volnou nabídku dostupných klíčových prvků, čeká netrpělivě rostlinný kořen, aby po „energetické a uhlíkové investici“ do půdy mohl pokrýt vlastní živinové nároky.
Shrnutí. Pro člověka je odjakživa velkým pokušením sklidit co největší úrodu, vykrmit co nejvíce dobytka. Nejsme připraveni na to, abychom pochopili, proč na vykrmení jednoho až dvou dobytčat na jednom hektaru musíme v půdě pastviny pro zachování její úrodnosti udržovat živoucí půdní havěť v souhrnném objemu až pětadvaceti dobytčat, z nichž je plně aktivní jen jedno. Nejsme připraveni na pochopení „paradoxu Šípkové Růženky“, na kouzlo „rhizosférního obchodování“ mezi živými partnery v půdě. Zato umíme nahradit jedinečnou půdní dovednost dodávat dusík, fosfor a další živiny nepřirozenými a ve svých důsledcích zhoubnými dodávkami minerálních hnojiv.
Ing. Jaroslav Záhora, CSc., (*1961) vystudoval Lesnickou fakultu Vysoké školy zemědělské v Brně. Disertační práci týkající se mikrobiálních transformací půdního dusíku obhájil v Ústavu systematické a ekologické biologie ČSAV, dále pak pracoval v oddělení lučních ekosystémů brněnské pobočky Botanického ústavu ČSAV. Krátce působil jako středoškolský učitel na soukromé škole pro zrakově postižené v Brně. V současnosti přednáší na agronomické fakultě Mendelovy univerzity v Brně. Zabývá se mikrobiální aktivitou půd.



ISSN 1213-0699 | ZO ČSOP Veronica | Panská 9, 602 00 Brno | mapa stránek časopisu